

As sementes são estruturas reprodutivas originadas após a fecundação, que consiste de um embrião com tecido de reserva (cotilédone ou endosperma) e envoltórios, e que é capaz de originar uma planta. Em sentido agronômico, denominamos as sementes, muitas vezes, como grãos. Os grãos nada mais são do que sementes que serão utilizadas para diversos fins, que não necessariamente para originar uma nova planta. No processo de formação da semente, a embriogênese compreende a formação do embrião, a partir da fertilização do óvulo (gameta feminino) pelo gameta masculino.
Primeiramente, vamos falar um pouco sobre sementes. Estas, compreendem a reprodução sexuada, que conferem variabilidade genética à espécie. Além disso, as sementes dão garantia à espécie de que haja a sobrevivência da mesma, uma vez que podem formar bancos de sementes que, posteriormente, darão origem a uma nova planta. Permitem, também, a colonização de outros ambientes. Nesse sentido, temos espécies com as mais diversas formas de dispersão de sementes, seja pelo vento, pela água, ou por animais. As plantas, dentro do seu processo evolutivo, desenvolveram diferentes maneiras de colonizar outros ambientes.
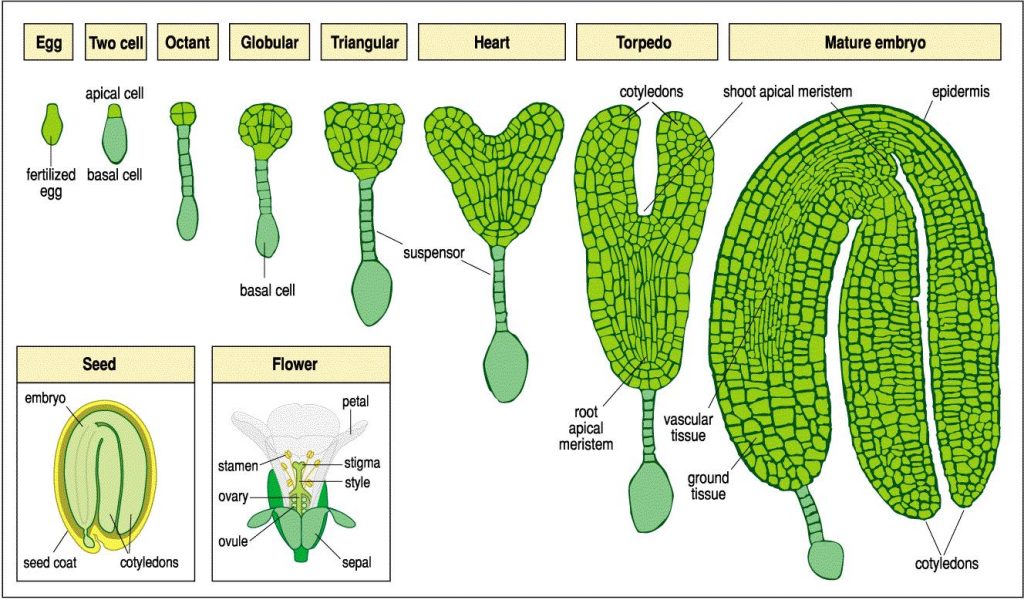
As sementes são formadas em plantas maduras, ou que já se reproduzem. Os grãos de pólen entram em contato com o estigma do pistilo, germinam e, a partir daí, ocorre a formação do embrião (embriogênese). O embrião é composto do eixo embrionário e pelos cotilédones. O eixo embrionário compreende o meristema apical do caule e da raiz. Em dicotiledôneas, ou plantas com dois cotilédones, o embrião passa basicamente por cinco estágios: zigótico, globular, de coração, torpedo e maduro. Note que, na Figura 1, há a presença de um “suspensor”, que é, basicamente, a ligação entre planta mãe e semente, que permite a nutrição para o embrião em desenvolvimento.
Estágio zigótico (two cell na Figura 1): compreende o primeiro estágio do ciclo de vida diploide (2n). Tem inicio na fusão da oosfera e do gameta masculino, formando um zigoto unicelular. Nesse estágio, ocorre o crescimento polarizado da célula (em uma direção apenas), seguido por uma divisão transversal, formando a célula apical e a célula basal.
Estágio globular (octant e globular na Figura 1): nesse estágio, a célula apical passa por uma série de divisões, gerando um embrião globular esférico de oito células (octante). Divisões celulares posteriores aumentam o numero de células do embrião globular, criando uma camada externa denominada de protoderme, que mais tarde se tornará a epiderme da semente.
Estágio de coração (heart na Figura 1): a partir deste estágio, ocorrem divisões celulares concentradas em duas regiões, ocorrendo ao lado do futuro meristema apical do caule, dando origem aos dois cotilédones (em dicotiledôneas).
Estágio maduro (mature embryo na Figura 1): esse estágio compreende o final da embriogênese, onde o embrião e a semente perdem água, tornando-se metabolicamente inativos na medida em que entram em dormência.
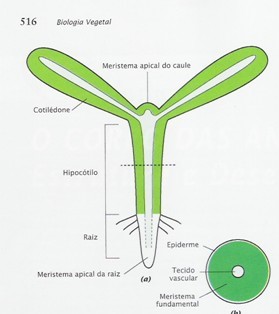
Há uma certa diferença na embriogênese de plantas dicotiledôneas e monocotiledôneas. Mudanças no tamanho, na forma, no número de células e nos padrões de divisão do embrião, são algumas destas diferenças, por exemplo. Ainda que ocorram algumas diferenças, de maneira geral, se pode dizer que existem muitas características em comum entre ambas, e que permitem serem generalizadas para todas as espermatófitas. A característica mais fundamental, nesse sentido, seja talvez a polaridade, onde se inicia com um zigoto unicelular, posteriormente, tornam-se mais polarizados pelo seu desenvolvimento ao longo de dois eixos: apical e basal, que compreende da extremidade do caule até a extremidade da raiz, e um eixo radial, que é perpendicular ao eixo apical-basal (Figura 2).

As sementes podem ser separadas, basicamente, em dois tipos: ortodoxas e recalcitrantes. As ortodoxas são aquelas sementes que passam pela dessecação da fase de maturação, enquanto as recalcitrantes não passam pela fase de dessecação que ocorre na maturação. Nesse sentido, as principais fases do desenvolvimento das sementes são:
- Alto conteúdo hídrico: divisão celular aumentada, aumento no peso fresco, histodiferenciação e morfogênese (formação do eixo embrionário, da polaridade, e do endosperma e cotilédones);
- Aumento da deposição de reservas: aumento do tamanho das sementes devido à expansão e ao acumulo de reservas (proteínas, lipídios e carboidratos). O tamanho do vacúolo é reduzido.
- Maturação: redução do metabolismo (quiescencia). Nas ortoxodas: dessecação. Nessa fase, de 90 a 95% do conteúdo de água é reduzido.
As sementes ortodoxas passam por um período de dessecação no estágio de maturação. A dessecação pode reduzir de 90-95% do conteúdo de água das sementes. Esta redução drástica faz com que a semente deva estar preparada para a ocorrência desta redução drástica no seu conteúdo de água. Nesse sentido, as proteínas LEA (Late embryogenesis abundant protein), ditas proteínas companheiras, tem grande importância na tolerância à dessecação pelas sementes. É o acumulo destas proteínas (LEA) que faz com que as sementes não sofram danos irreparáveis pelo decréscimo rápido no seu conteúdo de água. Estas proteínas que, acumuladas, dão suporte à semente no período de dessecação, são majoritariamente induzidas pelo Ácido Abscísico (ABA – Figura 3). As sementes recalcitrantes, por sua vez, associadas a ambientes mais úmidos, não passam pelo período de dessecação. Em termos agronômicos, as sementes ortodoxas tem maior viabilidade de armazenamento, por naturalmente terem baixa quantidade de água em sua composição. Enquanto isso, as sementes recalcitrantes necessitam maiores cuidados e tem “Shelf life” reduzido.
Os três principais hormônios envolvidos no desenvolvimento das sementes são: Ácido abscísico (ABA), Auxina (AIA) e Giberelina (GA). Nesse sentido, AIA e GA estão envolvidos com processos de divisão celular, ou a indução dos mesmos. Como explicamos anteriormente, a auxina está relacionada também com a polaridade na formação do embrião. Enquanto isso, o acúmulo de reservas, a tolerância à dessecação e a dormência estão associados com o ABA.
O ácido abscísico está envolvido no estimulo à dormência nas sementes. Isso faz com que tenha um papel muito importante ao evitar que as sementes germinem antes do tempo (viviparidade). Ou seja, o ácido abscísico previne que as sementes germinem antes mesmo de serem dispersas, dentro do fruto ou ainda na própria planta-mãe. Entramos, portanto, no tema de dormência de sementes.
Dormência de sementes:
Dormência é uma incapacidade temporária da semente de germinar em uma determinada condição ambiental que não impediria a germinação da semente não-dormente. Uma semente dormente não germina mesmo quando em condições ideais, devido a uma restrição interna ou sistêmica à germinação. O bloqueio pode ser na própria semente, ou na unidade de dispersão.
Uma semente quiscente, é uma semente que inicia e finaliza o processo germinativo quando da disponibilidade de condições ambientais favoráveis, e na ausência de elementos tóxicos. Esta é, portanto, a diferença entre uma semente dormente e uma quiescente. A semente em dormência não germina, porque há bloqueio interno, mesmo em condições ideais para germinação. Uma semente quiescente germina quando em condições ambientais ideais.
Podemos pensar na dormência como sendo um vilão: quando pensamos em produção em escala de alimentos. Ou podemos pensar na dormência como uma estratégia de sobrevivência: evolução das espécies. A dormência tem um papel biológico fundamental nas plantas. A dormência primária, que é aquela que vem com a semente, impede a germinação durante a fase final de maturação da semente. Além disso, a dormência ainda impede a germinação sincronizada após a dispersão das sementes, o que faz com que se reduza a competição pelas plantas, além de melhor distribuir espacial e temporalmente as sementes. A dormência ajuda, então, na distribuição das sementes no tempo, uma vez que elas irão depender da quebra da dormência.
Existem diversos mecanismos pelos quais as sementes podem tornar-se dormentes. Basicamente, a dormência pode ser imposta pelo embrião, ou pelos envoltórios. Quando a dormência é imposta pelo embrião, o mesmo pode estar indiferenciado, ser imaturo, ou conter inibidores químicos ou restrições fisiológicas à germinação. A dormência imposta pelos envoltórios tem, normalmente, interferência na absorção de água, nas trocas gasosas, impedimento da saída de inibidores ou restrições mecânicas. A diferença aqui é que a imposição da dormência pelos envoltórios significa, basicamente, uma restrição mecânica ao processo de germinação. Por exemplo: tegumento rígido ou impermeável, que não permite a passagem de água e/ou a saída de componentes que mantenham a dormência da semente.
Em relação a dormência imposta pelo embrião, embriões indiferenciados ainda não sofreram histodiferenciação, e não passou pela fase de maturação. Embriões imaturos, precisam terminar a fase de maturação antes de germinar. Além disso, podem haver também inibidores químicos que impedem a germinação por impedir o crescimento do embrião. Estes, não estão relacionados com hormônios vegetais, como o Ácido Abscísico. Podem ser, por exemplo, a ocorrência de oxidação de substancias que acabam por reduzir o oxigênio do meio.
Como dito, então, há influência dos hormônios vegetais na dormência das sementes. Nesse sentido, o controle hormonal da dormência das sementes inclui, basicamente, a concentração de dois hormônios: ABA e GA. Quando as concentrações de ABA são maiores que as concentrações de GA, a dormência prevalece. Quando as concentrações de GA são maiores do que as concentrações de ABA, a germinação é induzida. Nesse caso, o que ocorre é que a semente dormente tem seus níveis de ABA diminuídos, assim como sensibilidade ao ABA, ao mesmo tempo em que se aumentam os níveis de GA e/ou da sensibilidade ao mesmo. Estes eventos induzem a ruptura da testa. Posteriormente, com ação de GA e Brassinosteroides (hormônios que também participam da germinação), agindo juntamente na ruptura do endosperma, a semente consegue germinar. Nesta etapa, o ABA pode agir de maneira oposta, bloqueando a ruptura do endosperma, ou a germinação em si. O etileno também pode participar deste processo bloqueando a ação do ABA.
Em resumo: a quebra da dormência é depentende de uma mistura de três fatores:
- Catabolismo do ABA;
- Menor sensibilidade ao ABA;
- Maior concentração de GA.
Há, ainda, um outro conceito de dormência. A dormência cíclica ocorre em casos em que as sementes passam por diversas condições ambientais até que cheguem a condição ambiental ideal que as permita germinar completamente. Nesse caso, as sementes, expostas a ambientes favoráveis, perdem a dormência e, posteriormente, quando retornam a condição não-favorável, se tornam dormentes novamente até ter as condições ideais. A dormência é cíclica, nesse caso, pois passam por ciclos de dormência e sem dormência, por diversas vezes.
As espécies reativas de oxigênio (ROS) podem também influenciar na dormência das sementes. Por exemplo, o ABA induz a síntese de enzimas antioxidantes (CAT2), que bloqueiam a síntese de ROS, reduzindo os níveis de espécies reativas de oxigênio. Portanto, a catalise de ROS induzida pelo ABA, realizada pela enzima catalase (CAT2), afeta a germinação na medida em que as ROS são sinalizadoras de síntese de GA, fazendo com que seja reduzida a biossíntese de GA. Por outro lado, a presença de ROS induz a síntese de enzimas que fazem o catabolismo do ABA (reduzindo os seus níveis), induzindo, por duas vias, a germinação das sementes.
Fontes utilizadas:
ISHIBASHI, Yushi et al. The interrelationship between abscisic acid and reactive oxygen species plays a key role in barley seed dormancy and germination. Frontiers in plant science, v. 8, p. 275, 2017.
TAIZ, Lincoln et al. Fisiologia e desenvolvimento vegetal. Artmed Editora, 2017.
FINCH‐SAVAGE, William E.; LEUBNER‐METZGER, Gerhard. Seed dormancy and the control of germination. New phytologist, v. 171, n. 3, p. 501-523, 2006.
BEWLEY, J. Derek et al. Seeds: physiology of development, germination and dormancy. Springer Science & Business Media, 2012.

