

A fase fotoquímica – ou reações luminosas – consiste no primeiro conjunto de reações que englobam a fotossíntese e que ocorrem na membrana do tilacóide. Como o próprio nome já diz e você já sabe ou pode imaginar, são reações que envolvem a luz solar. Já adianto, ao final de todo o processo serão produzidos compostos de alta energia: ATP e NADPH. O que acontece é que a energia luminosa é convertida em energia química por duas unidades funcionais conhecidas como fotossistemas. Essa energia da luz serve como um “gatilho” que impulsiona a transferência de elétrons por uma série de compostos que podem ser receptores ou doadores. A maior parte desses elétrons é extraída da H20 – que é oxidada a O2. Além disso, a energia luminosa também apresenta papel fundamental na força motriz de prótons – H+ – que é utilizada para formar ATP. Até agora foi apenas uma introdução, vamos então ver como tudo isso acontece passo a passo.
O primeiro ponto a ser entendido é que não existe fotossíntese sem absorção da luz solar e não existe absorção sem pigmentos. A planta possui pigmentos que estão relacionados diretamente com a fotossíntese, sendo então conhecidos como pigmentos fotossintetizantes. Os principais são as clorofilas, que apresentam uma estrutura complexa em forma de anel com uma longa cauda de hidrocarbonetos quase sempre ligada à essa estrutura. Nas plantas são comumente encontradas as clorofilas a e b.
A clorofila absorve – principalmente – a luz vermelha (625 – 740 nm) e azul (440 – 485 nm) do espectro, sendo que a luz verde – cerca de 550 nm – é refletida, por isso enxergamos a folha verde. Os carotenoides são outros pigmentos encontrados nos organismos fotossintetizantes. Esses pigmentos são conhecidos como pigmentos acessórios devido toda energia absorvida por eles ser transferida para as clorofilas. Além disso, desempenham papel importante na defesa do organismo de danos causados pela luz. Os carotenoides absorvem luz na região dos 400 – 500 nm, resultando em sua coloração alaranjada (Fig. 1).

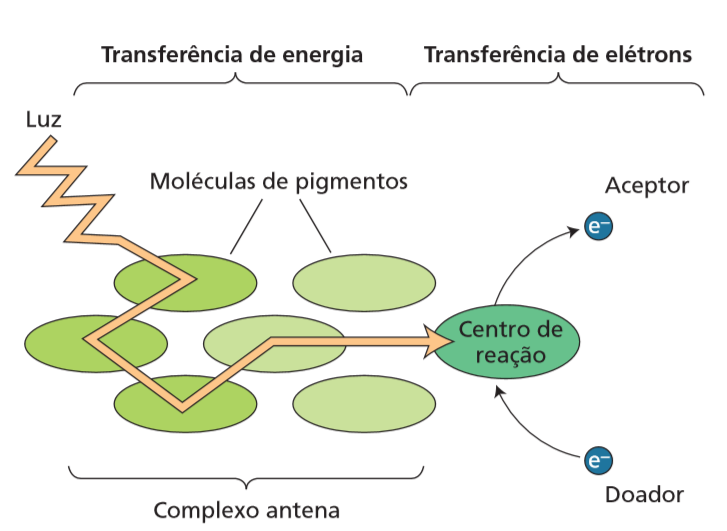
Agora vamos entender como ocorrem as reações da etapa fotoquímica. Parte da luz absorvida pelos pigmentos fotossintetizantes – clorofilas e carotenoides – é armazenada como energia química. A maior parte dos pigmentos serve como complexo antena que capta a luz e transfere a energia por vibração até chegar no centro de reação (Fig. 2). Vocês podem se perguntar: Por que existe essa divisão entre complexo antena e centro de reação ? É exatamente porque mesmo sob grande incidência luminosa a clorofila consegue absorver poucos fótons e se apenas uma clorofila estivesse ligada ao centro de reação, as enzimas desse centro estariam ociosas a maior parte do tempo.
O esquema Z (zigue-zague) tornou-se a base para compreensão de como ocorre toda essa transferência de elétrons e consiste em dois fotossistemas – PSI e PSII – física e quimicamente diferentes (Fig. 3A). O PSI absorve preferencialmente a luz vermelho – distante de comprimentos maiores que 680 nm, enquanto o PSII absorve luz vermelha de comprimento de 680 nm. O PSI produz um redutor forte – reduz NADP+ – e o PSII produz um oxidante forte – oxida a água.
Quando a luz vermelha incide no PSII, as clorofilas se excitam e transferem toda energia por vibração para o centro de ração P680 (oxidante forte). A clorofila P680 transfere 1 é para feofitina (pheo).
Quem repõe os elétrons no PSII é a H20. Sempre ocorre a oxidação de duas moléculas de H20 para que não ocorra a parcialidade da oxidação e não promova a formação de espécies reativas de oxigênio. Quem é responsável por isso é o complexo de evolução do oxigênio, liberando com a quebra da molécula de água 4 elétrons. Mas claro que esses elétrons também não podem ser liberados de uma vez no PSII, caso contrário a chance de formas ROS seria alta. Dessa forma, íons de manganês e cálcio, que estão presentes nesse complexo, “armazenam” esses elétrons e liberam um por vez.
A pheo transfere o elétron para as plastoquinonas PQa e PQb. O complexo citocromo b6f (citb6f) transfere o elétron recebido da PQ para a plastocianina (PC), que por sua vez reduz o P700.
A PQb, que é o último receptor do PSII, recebe um elétron e fica em um estado semi reduzido, isso porque ela comporta dois elétrons e não apenas um, onde ela permanece enquanto aguarda o segundo. Quando recebe o segundo elétron vindo pelo mesmo caminho, fica totalmente reduzida e passa a ser chamada de plastoquinol. Esse transportador se desliga do PSII e migra pela membrana do tilacoide, já que é hidrofóbico, até chegar no citb6f, onde descarrega seus elétrons.
A PQH2 transfere os elétrons para o citb6f que por sua vez transfere para plastocianina (PC). O problema é que o citocromo recebe dois elétrons e a plastocianina tem capacidade de receber apenas um. Assim, um dos dois elétrons que chega no citb6f percorre o caminho mais rápido para a PC e o outro migra por um caminho por dentro do citocromo até o segundo sítio de ligação da plastoquinona, tendo agora uma plastoquinona semi-reduzida (Fig. 3). Lembrando que ela recebe dois elétrons, permanece assim até a chegada de um segundo, reciclando assim os elétrons e evitando perdas. Com isso, dois prótons são retirados do estroma para o lúmen.
Após ser reduzido pela PC, o PSI transfere seus elétrons para os aceptores de elétrons seguintes, A0 e A1, que são uma clorofila e uma quinona respectivamente. Os elétrons são então transferidos para uma série de proteínas ferro-sulfurosas ligadas a membrana – FeSX, FeSA, FeSB – que logo em seguida transfere os elétrons para ferredoxina (Fd). A flavo-proteína solúvel ferredoxina- NADP+– redutase (FNR) reduz o NADP+ a NADPH, que é utilizado no ciclo de calvin.
Os prótons que estão agora no lúmen retornam para o estroma passando pela ATPsintase, ativando-a e gerando ATP (Fig. 3B).
Além de tudo isso, há ainda a fosforilação cíclica, que consiste em uma adaptação exclusiva para uma maior demanda de ATP e não para formação de NADPH. Nesse tipo de fosforilação, a ferredoxina é substrato para outra enzima, a ferredoxina-plastoquinona oxiredutase, que reduz a PQ a PQH2, por meio da transferência dos elétrons da Fd de volta para a PQ. Dessa forma, mais prótons são transportados para o lúmen, colaborando assim para uma maior produção de ATP. No entanto, como os elétrons que a Fd transferiria para o NADP+ foram para a PQ, não há formação de NADPH.

Referências utilizadas
Taiz, L., Zeiger, E., Møller, I. M., & Murphy, A. Fisiologia e desenvolvimento vegetal. Artmed Editora (2017)
BUCHANAN, Bob B.; GRUISSEM, Wilhelm; JONES, Russell L. (Ed.). Biochemistry and molecular biology of plants. John Wiley & Sons (2015)

